
POTENCIALES DE MEMBRANA
|
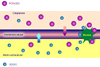
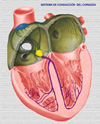
Las células cardíacas, como las otros tejidos excitables de los mamíferos, tienen una composición iónica intracelular que difiere de la extracelular. La concentración de iones potasio K+ en el interior de la célula es unas 30 veces mayor que concentración extracelular, mientras que el sodio Na+ es unas 30 veces menor. Como la membrana de las células cardíacas es más permeable al potasio que al sodio, en condiciones de reposo, los iones de potasio pueden salir de la célula con más facilidad que pueden entrar los iones de sodio y, en consecuencia existe una diferencia de potencial entre el interior y el exterior de la célula. Se dice que la membrana de la célula está polarizada, siendo la diferencia de potencial de - 90 mV (esto quiere decir que hay más cargas negativas en el interior que el exterior) Cuando un estímulo llega a una de estas células, se alteran las propiedades fisicoquímicas de la membrana, aumentando su permeabilidad al sodio. Como el sodio está mucho más concentrado en el exterior de la célula (se dice que existe un gradiente de concentración), al aumentar la permeabilidad de la membrana, el sodio entra de forma masiva en el interior de la célula. Esta afluencia del cargas positivas invierte el potencial de membrana. Si se registra en un osciloscopio la secuencia de acontecimientos que tienen lugar cuando se estimula una célula cardiaca, en condiciones basales, el aparato registra un voltage de - 90 mV que se mantiene estable en tanto que el estímulo no llega a la célula. El cambio súbito de permeabilidad de la membrana cuando llega el estímulo, con la entrada masiva de iones sodio en la célula, se refleja en aumento muy rápido del voltaje a +20 mV (fase 0 o fase de excitación). Después del período de excitación, hay un período de duración variable durante el cual el potencial es prácticamente 0 (meseta del potencial o fase 2). Seguidamente, se produce un nuevo cambio de la permeabilidad de la membrana que, en este momento se hace más permeable al potasio, que sale rápidamente de la célula, restaurándose el potencial a los niveles anteriores a la llegada del estímulo (fase de repolarización o fase 3). Sigue un período estable de reposo, hasta la llegada de un nuevo estímulo (fase 4). Los cambios registrados en el osciloscopio desde la llegada del estímulo hasta la vuelta al estado de reposo reciben el nombre de potencial de acción (*) No todas las células cardiacas tienen los mismos potenciales de acción: Nodo sinusal: este es un pequeño nódulo (también llamado marcapasos), situado en la aurícula derecha, bajo la desembocadura de la vena cava (*). El registro osciloscópico de los potenciales de las células de este nódulo muestras dos importantes características:
Este comportamiento explica el automatismo de las células del marcapasos. No es necesaria la llegada de un estímulo para provocar el cambio de la permeabilidad de la membrana a los iones, sino que dicha permeabilidad al Na+ primero y al K+ se instaura espontánea y ciclicamente a un ritmo de 60 a 100 veces por minuto. Nodo auriculo-ventricular: los potenciales de acción registrados en las fibras del nodo A-V son muy parecidas a las que se presentan en el nodo sinusal (*) Sistema de His-Purkinje: los potenciales de acción de las fibras de esta parte del sistema de conducción tienen tres propiedades importantes:
Las demás células cardíacas muestran potenciales de acción intermedios entre los de seno sinusal y las fibras de Purkinje (*) Existe un cierto decalaje temporal entre el estímulo eléctrico y la respuesta contráctil de las fibras musculares (*). La tensión muscular máxima tiene lugar al final de la repolarización, siguiendo rapidamente la relajación |
|
Potenciales rápidos y potenciales lentos (*) La despolarización de la membrana de las células cardíacas depende de la naturaleza de las células. En el nodo sinusal y en el nodo auriculoventricular, esta fase del potencial de acción es lenta mientras que en las células del sistema His-Purkinje es muy rápida. Esto se debe a la diferente permeabilidad de las membranas a los iones, permeabilidad condicionada, a su vez, por la presencia de los canales iónicos. Los canales iónicos están constituidos por unas proteínas transmembrana que tienen la propiedad de abrirse o cerrarse para dejar pasar o no, determinados iones. En las células cardíacas existen cuatro tipos de canales selectivos para sodio, potasio, calcio y cloro. A efectos del comportamiento de los potenciales de acción, los más importantes son los tres primeros. Los potenciales rápidos se encuentran en las células ventriculares y en las del sistema His-Purkinje. Se caracterizan por:
Los potenciales lentos se localizan en las células del nodo sinusal y el nodo aurículo ventricular. Se caracterizan por:
La despolarización se lleva a cabo mediante la entrada de sodio y de calcio a través de sus correspondientes canales lentos. En estas células no hay canales de sodio rápidos Estas características contribuyen a una conducción más lenta de estos potenciales y a un periodo refractario mayor en los tejidos que muestran unas respuestas lentas. |
 |
Se denominan períodos refractarios los tiempos del ciclo de excitación de una célula cardíaca durante los cuales un nuevo estimulo no produce ninguna respuesta por no haberse completado los ciclos de apertura/cierre de las puertas de los canales. Esto ocurre durante las fases 0, 1, 2 y parte de la 3 (*) y explica porque no puede haber una contracción hasta que la membrana celular no se ha recuperado del estímulo anterior. Por la misma razón, las fibras cardíacas no pueden tetanizarse. Además del período refractario absoluto, existe el período refractario relativo, al final de la fase de repolarizacíón, durante el cual si es posible despolarizar nuevamente la célula siempre y cuando el estímulo sea lo suficientemente intenso. Como es lógico, la duración de los períodos refractarios está directamente relacionada con la duración del ciclo de excitación. Esta relación permite explicar el fenómeno de Ashman: la súbita prolongación de la duración de un ciclo prolonga el perìodo refractario para el siguiente impulso que, si llega demasiado pronto, se encuentra con un tejido refractario, produciendo un ralentizamiento de la conducción en esta área. Los periodos refractarios sin embargo, no son solo afectados por la frecuencia de la estimulación sino también por factores que influyen sobre el medio iónico, fármacos y estados patológicos como la isquemia o la hipoxia |




   |